Abstract
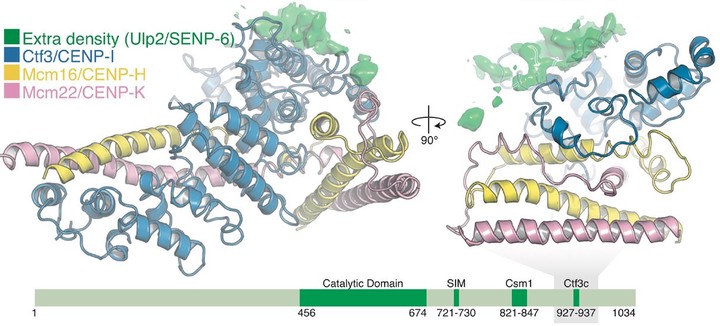
The step-by-step process of chromosome segregation defines the stages of the cell cycle. In eukaryotes, signals controlling these steps converge upon the kinetochore, a multiprotein assembly that connects spindle microtubules to chromosomal centromeres. Kinetochores control and adapt to major chromosomal transactions, including replication of centromeric DNA, biorientation of sister centromeres on the metaphase spindle, and transit of sister chromatids into daughter cells during anaphase. Although the mechanisms that ensure tight microtubule coupling at anaphase are at least partly understood, kinetochore adaptations that support other cell cycle transitions are not. We report here a mechanism that enables regulated control of kinetochore sumoylation. A conserved surface of the Ctf3/CENP-I kinetochore protein provides a binding site for Ulp2, the nuclear enzyme that removes SUMO chains from modified substrates. Ctf3 mutations that disable Ulp2 recruitment cause elevated inner kinetochore sumoylation and defective chromosome segregation. The location of the site within the assembled kinetochore suggests coordination between sumoylation and other cell cycle–regulated processes.